PRTF - Perry Rhodan Technik Forum
Grundlegende Gedanken zur Biochemie
und
molekularen Genetik im Perryversum
(c) Michael Gebinoga 15. 11. 1999
Einführung
Ein Punkt, der bei PR in einem hohen Maße hineinspielt, ist die biochemische Kompatibilität bzw. Inkompatibilität von verschiedenen Lebensformen und die Entstehung des Lebens als solches. Wie können wir uns derartiges vorstellen? Im Perryversum werden bestimmte Randbedingungen erwähnt, die einem einen gewissen Anhaltspunkt geben, jedoch werden einem keine genauen Spezifikationen an die Hand gegeben. Wir sind also auch auf diesem Gebiet auf Annahmen angewiesen. Einige Themen im Perryversum, über die man unter diesen Aspekten diskutieren könnte, wären wie folgt:
- Biochemie der sauerstoffatmenden Lebewesen im Perryversum
- Aufbau des Virenimperiums der Kosmokratin Vishna
- Funktionsweise eines Zellaktivators
- Taurecs genetische Experimente in Truillau und insbesondere die Erschaffung von Voltago
- Eigenschaften der On und Noon Quanten in den Sporenschiffen der Mächtigen
- Genetische Experimente von Monos in der Milchstraße
- Kybernetisch-biologische Modifikationen der Cantaros
- Medizin der Aras und kosmomedizinische Zentren in der Milchstraße (z. B. Mimas, Tahun, Aralon)
- Konvertermagen und Strukturvariabilität der Haluter bzw. die genetischen Experimente der Konstrukteure des Zentrums in M87 (Druithora)
- Kompaktkonstitution der Oxtorner
- Biochemie der Maahks
Ich werde versuchen, im Lauf der Zeit auf einige der angesprochenen Punkte detailliert einzugehen, jedoch sind vor einer derartigen Betrachtung ein paar grundlegende Überlegungen notwendig. Wenn wir fürs erste Lebewesen ausklammern, die einen völlig fremdartigen Metabolismus haben (z.B. Maahks oder energetische Lebensformen), können wir uns auf die Lebensformen beschränken, die auch unter irdischen Bedingungen lebensfähig wären. Dazu zählen neben der Notwendigkeit von Sauerstoff für die Atmung, der auf Kohlenstoff basierenden Biochemie noch folgende molekularbiologische Details:
- Identische Chiralität (Händigkeit) von Aminosäuren als Proteinbausteine und den gängigen Nucleotiden als Nucleinsäurebausteine.
- DNA als Erbsubstanz und ähnlicher genetischer Code.
Chiralität
Als Chiralität bezeichnet man die Eigenschaft von makroskopischen Dingen und Molekülen, als Bild und Spiegelbild vorliegen zu können. Beispiele aus dem täglichen Leben sind unsere Hände, aber auch solche Dinge wie ein Korkenzieher, eine Schraube oder ein Schneckenhaus. Bei Molekülen ist diese Eigenschaft meistens eine direkte Folge der auf dem vierbindigen Kohlenstoff basierenden Biochemie. Hat ein Kohlenstoffatom (C) vier unterschiedliche Atomgruppen gebunden, so haben wir ein chirales Molekül vorliegen. Ein einfaches Beispiel für Chiralität sind die rechte und die linke Hand des Menschen. Obwohl sie beide identisch aussehen, kann man sie doch nicht zur Deckung bringen, oder anders gesprochen, man kann mit der rechten Hand nicht in einen linken Handschuh hineinschlüpfen. Beispiele aus der Biochemie sind Milchsäure und Ethanol. Bei Milchsäure haben wir ein chirales Molekül vorliegen, wohingegen Ethanol nicht chiral ist.
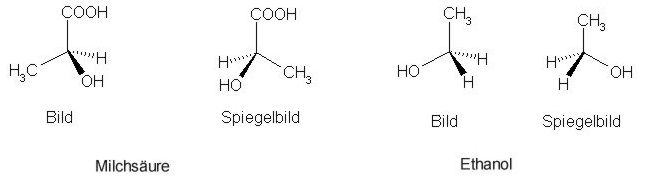
In dem Bild sind die durch Striche gekennzeichneten Bindungen als in der Papierebene vorliegende Bindungen anzusehen. Die vier Bindungen laufen im Zentrum auf ein C-Atom zusammen, welches in dieser Darstellungsweise nicht eingezeichnet ist. Die gestrichelten Bindungslinien gehen nach hinten weg und die als schmale Keile vorliegenden Bindungen kommen aus der Papierebene nach vorne heraus. Wir sehen also, dass wir durch diese Darstellung Moleküle als dreidimensionales Bild und Spiegelbild zeigen können. Wenn wir nun die jeweiligen Bilder von Milchsäure und Ethanol um die nach oben weisende Bindungsachse drehen, sehen wir, dass wir Bild und Spiegelbild beim Ethanol zur Deckung bekommen können. Ethanol ist achiral. Bei der Milchsäure hingegen ist das nicht möglich. Genausowenig könnten wir einen rechten und einen linken Handschuh ineinander stecken. Wir haben bei der Milchsäure folglich ein chirales Molekül vorliegen. Chirale Moleküle haben ansonsten vollkommen identische Eigenschaften, wie Schmelzpunkt, Löslichkeit und Dichte. Erst wenn ein anderes chirales Medium mit ihnen wechselwirkt, offenbaren sie ihre chirale Natur. Im Falle der Aminosäuren ist das sowohl linear polarisiertes Licht als auch Enzyme. Bei linear polarisiertem Licht wird die eine Ebene des Lichts stärker gedreht als die andere, abhängig wieder davon, ob wir eine Lösung mit dem Bild oder dem Spiegelbild des Moleküls durchstrahlen. Man spricht bei diesen optisch aktiven Substanzen auch von rechtsdrehender oder linksdrehender Milchsäure ((+) Milchsäure und (-) Milchsäure). Neben den Aminosäuren ist normaler Zucker ebenfalls optisch aktiv. Enzyme sind wiederum ebenfalls aus chiralen Aminosäuren aufgebaut und können beispielsweise nur das Bild oder das Spiegelbild eines Moleküls verarbeiten.
Alle biologisch wichtigen Moleküle unterliegen dem chiralen Prinzip, angefangen von den Bausteinen biologischer Makromoleküle, den Aminosäuren und Nucleotiden, bis zu den daraus resultierenden Makromolekülen, den Proteinen (Polyaminosäureketten) und den Nucleinsäuren (Polynucleotidketten). Biologische Aminosäuren sind allesamt linksdrehend, d. h., wenn man eine wässrige Lösung einer natürlichen Aminosäure mit polarisiertem Licht bestrahlt, wird die Ebene des Lichts um einen kleinen Betrag nach links gedreht. Rechtsdrehende Aminosäuren können von einem Organismus im allgemeinen nicht verarbeitet werden, wenn man von einigen seltenen Ausnahmen absieht. Zwei weitere Beispiele über chirale Moleküle und ihre Wirkung auf den Menschen sind Adrenalin und Contergan. Während (-)-Adrenalin den Herzmuskel anregt, ist die (+)-Komponente völlig wirkungslos. Bei Contergan ist der Unterschied noch gravierender. Die eine Komponente ist ein wirkungsvolles Schlafmittel ohne Nebenwirkung, während die andere Komponente starke Schädigungen an ungeborenen Embryonen hervorruft. Hierzu ist noch zu sagen, dass bei einer chemischen Synthese meistens beide Komponenten in gleicher Menge gebildet werden. Die chirale Trennung in die jeweiligen Komponenten ist ein ausgesprochen aufwendiger Prozeß.
Würden einem Mensch beispielsweise nur Lebensmittel aus rechtsdrehenden Aminosäuren zur Verfügung stehen, würde dieser Mensch buchstäblich verhungern, auch wenn er von der Menge her, genug Nahrung zur Verfügung hätte. Dies ist auch zwingend notwendig, wenn man sich den Genüssen der gatasischen Küche zuwenden will. Die Tatsache, dass es bestimmte Stoffe gibt, die für Lemurabkömmlinge (Terraner, Akonen, Arkoniden, Tefroder etc.) giftig sind, während z. B. Blues diese problemlos zu sich nehmen können, wird hiervon nicht berührt.
Es wäre zu überlegen, in welcher Weise man für unser Standarduniversum neben dem Strangenessfaktor ein chirales Prinzip einführen kann. Dieses Prinzip würde besagen, dass in Lebewesen vorkommende chirale Bausteine (z. B. Aminosäuren) dieselbe Chiralität aufweisen würden. Dies ließe sich durch mehrere Mechanismen realisieren, die alle ihre Vorteile und Nachteile haben.
Zum einen könnte man eine entsprechende Eigenschaft der On und Noon Quanten postulieren. Dadurch wäre gewährleistet, dass sämtliches aus diesen Vorstufen stammendes Leben prinzipiell miteinander kompatibel wäre. Genetische Verschmelzungen wie die von Taurec in Truillau durchgeführten genetischen Experimente wären dadurch überhaupt erst möglich. Andererseits könnte man bei diesem Mechanismus auch andere Lebensformen einführen, die nicht aus On und Noon Quanten entstanden sind und beispielsweise eine andere Chiralität aufweisen könnten.
Im anderen Fall würde man die biologisch richtige Chiralität als inhärente Eigenschaft unseres Universums einführen. Kommt es zu einem Transfer von einem Universum in ein anderes würde sich die Chiralität automatisch anpassen, vergleichbar der Strangeness-Anpassung bei Inter-Universums-Reisen. Diese Sichtweise würde beispielsweise erklären, warum Perry Rhodan das Essen der Hauri bei seinem Aufenthalt im Tarkan Universum zwar nicht schmeckte, aber durchaus verträglich war. Das ein Zellaktivator Giftstoffe umwandeln kann, wird zwar vorausgesetzt, jedoch ist es wenig wahrscheinlich, dass ein ZA die Chiralität von Aminosäuren ändern kann. Das gleiche gilt selbstverständlich für den Metabolismus der Kartanin bei ihrem Wechsel vom Tarkan-Universum ins Standard-Universum.
Bezüglich der Chiralität möchte ich auch noch die Kritik von Michael Deutsch einfügen, dem die Chiralität als inhärente Eigenschaft unseres Universums nicht behagte. Zitat: "Eine universelle Konstante, die sich beim Transfer anpaßt? In einem anderen Universum können nach der Hyperabtraktumholistik entweder gleiche, ähnliche oder völlig andere Kausalitäten herrschen. Bei einem Transfer würde sich also der tranferierte Organismus anpassen (oder ziemlich schnell sterben), nicht aber seine universellen Konstanten."
Nach einiger Überlegung möchte ich Michael Deutsch hinsichtlich seiner Kritik zustimmen. Darüberhinaus bieten verschiedene Chiralitäten im Standarduniversum dramaturgisch mehr Möglichkeiten. Zu der Überlegung von chiralitätsdeterminierenden On und Noon Quanten sowie Taurecs Experimenten ebenfalls nochmal Michael Deutsch's Kommentar: "Eine mögliche Erklärung, aber eigentlich nicht nötig. Bei seinen (Taurecs) Experimenten mußte er eh auf molekularer und submolekularer Ebene agieren, warum also nicht gleich die Chiralität determinieren? Aber der Gedanke an eine Nicht-O/NO-Q-Lebensform ist schon interessant... Vielleicht bevorzugen ja die Chaotarchen sowas?"
Bezüglich der On/Noon Quanten würde ich vorschlagen, eine chirale Determinierung zu postulieren. Die bisher in die Serie eingebrachten Lebensformen und ihre Interaktionen mit fremden Ökosphären zeigen klar auf, dass es bisher keine diesbezüglichen Unverträglichkeiten gab. Dadurch ließen sich auch die Aspekte der nicht auf On/Noon Quanten beruhenden Lebensformen geschickt in die Serie einbinden. Dies müßten nicht unbedingt besonders exotische Lebensformen sein, allein ihre auf völlig verschiedenen Grundprinzipien beruhende Biochemie würde mannigfaltige Möglichkeiten eröffnen.
Erbsubstanz DNA und genetischer Code
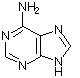
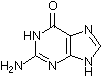
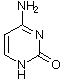
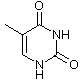
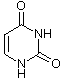
Obwohl es inzwischen einige andere Möglichkeiten gibt, das Erbgut in Form eines replikationsfähigen Trägermoleküls zu realisieren, sollte man aus praktischen Erwägungen auch für die gängigen sauerstoffatmenden Lebewesen DNA (Desoxyribonucleinsäure) als Träger des Erbguts annehmen. DNA ist stabil und bietet eine hinreichende Menge an Möglichkeiten, verschiedenste Lebensformen zu realisieren, wie es die Evolution über Jahrmillionen auf der Erde gezeigt hat. Die DNA setzt sich dabei aus einzelnen Desoxynucleotiden zusammen. Die Nucleotide setzen sich aus einer stickstoffhaltigen Base, einem Zucker (Pentose) und einer Phosphatgruppe zusammen. Die stickstoffhaltigen Basen (Adenin, Guanin, Cytosin und Thymin) definieren dabei in ihrer Abfolge die Erbinformation, wohingegen Zucker und Phosphatgruppe das Rückgrat der Polynucleotidkette bilden. Auch hinsichtlich der zugrundeliegenden Bausteine wie den Pentosezuckern sowie den Nucleotidbasen liegen chirale Bedingungen vor. Eine weitere wichtige Nucleinsäure ist RNA, welche ein paar kleine Unterschiede zur DNA aufweist. Bei der RNA wird z. B. statt Thymin Uracil verwendet.
|
Adenin |
Guanin |
Cytosin |
Thymin (nur bei DNA) |
Uracil (nur bei RNA) |
|
|
|
|
|
|
Im Erbgut selbst, liegt ein Strang DNA mit einem zweiten,
antiparallelen und komplementären DNA Strang gemeinsam vor.
Die Komplementarität ergibt sich aus der "Passform“ von
Adenin und Thymin (A und T) zueinander, sowie von Cytosin und
Guanin (C und G) zueinander. Ein kurzes Gen auf einem DNA-Strang
kann z. B. wie folgt gelesen werden:
ATG GCG CTA TGG TAA ("sense" Strang)
TAC CGC GAT ACC ATT (Komplementärer Gegenstrang;
"antisense" Strang)
Dabei ist zu bemerken, dass der Gegenstrang von rechts nach links gelesen wird, also TTA CCA... etc. Des weiteren werden die einzelnen Nucleotide durch ihre Nucleotidbasen (A, C, G und T) abgekürzt wiedergegeben.
Dieselbe Überlegung gilt für den genetischen Code. Wir
haben vier verschiedene Nucleotidbausteine und 20 verschiedene
Aminosäuren, aus denen sich Nucleinsäuren und
verschiedenste Proteine aufbauen. Dabei kodieren jeweils drei
Nucleotidbausteine (Tripletts) für eine Aminosäure. Diese
Zuordnungstabelle von Nucleotidtripletts (Codons) und
Aminosäuren nennt man den genetischen Code. Wir haben also
auch für die Mehrzahl der Lebewesen einen
übereinstimmenden genetischen Code. In dem obigen Beispiel des
kurzen DNA-Strangs würden wir die genetische Botschaft wie
folgt lesen:
| DNA Ebene | ATG | GCG | CTA | TGG | TAA |
| Protein Ebene | Met | Ala | Leu | Trp | Stopp |
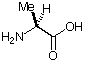
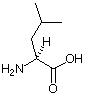
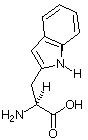
| Aminosäurestruktur | 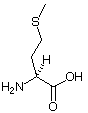 |
 |
 |
 |
- |
Die Aminosäureabkürzungen lauten: Met = Methionin, Ala = Alanin, Leu = Leucin, Trp = Tryptophan.
ATG ist in den meisten Fällen das erste Triplett in einem Gen und damit Methionin die erste Aminosäure im frisch synthetisierten Protein. TAA ist eines der drei Stoppcodons, welche angeben, dass an dieser Stelle das Gen endet. Die Stoppcodons haben dabei keine entsprechende Aminosäure. Der normale genetische Code ist hochgradig redundant, da es bei Nucleotidtripletts aus insgesamt vier Nucleotidbausteinen 64 Möglichkeiten gibt, diese anzuordnen. Demgegenüber haben wir aber nur 20 Aminosäuren. D. h., mehrere Codons kodieren für ein und dieselbe Aminosäure. Bei einigen Aminosäuren, z. B. Arginin (Arg) sind es sogar sechs Codons, die für dieselbe Aminosäure kodieren. Würden wir von einem genetischen Code ausgehen, der noch zwei zusätzliche Nucleotidbausteine verwendet, würde die Zahl der möglicher Codons auf 216 anwachsen.
Diese Redundanz des genetischen Codes erlaubt es uns im Perryversum zusätzliche Aminosäuren zu nutzen, die Proteinen neue Eigenschaften verleihen können. Dies ist von besonderem Interesse bei einigen genetischen Experimenten und bei der molekularbiologischen Deutung von Oxtornern, Halutern und weiteren Lebewesen mit Extremkonstitution. Bei diesen Lebewesen kann man auch überlegen, ob es Sinn macht einen erweiterten genetischen Code zu verwenden (6 Nucleotidbausteine, 216 mögliche Codons), um eine Vielzahl von verschiedenen Aminosäuren zu nutzen. Darüberhinaus könnte ein solcher Code als relativ wenig anfällig gegen Mutationen designt werden. Bei sechs Nucleotidbausteinen hätte man bereits 36 Zweierkombinationen vorliegen und folglich würde das 35 mögliche Aminosäuren bedeuten und eine Stoppposition. Bei diesen Kombinationen wäre die dritte Stelle folglich nicht relevant. Im normalen genetischen Code kommt dies bereits ebenfalls vor.
Aminosäure Codons
Prolin (Pro) CCU, CCC, CCA, CCG
Jedoch auch ohne eine Erweiterung bietet der genetische Code
eine Menge Spielraum, sowohl für Erweiterungen, als auch
für die Verwendung von neuen Aminosäuren mit
ungewöhnlichen Eigenschaften. Daraus folgen würden dann
gleichfalls Proteine mit entsprechenden Eigenschaften. Man darf
nicht vergessen, dass bereits die auf dem normalen genetischen Code
beruhende Lebensvielfalt überwältigend groß ist.
Würde man nur versuchen, alle möglichen Proteine mit
einer Kettenlänge von 200 Aminosäuren herzustellen (es
gibt auch kleinere und deutlich größere Proteine), dann
würde bereits ein winziger Bruchteil davon ausreichen, um das
gesamte Universum dicht und homogen mit Proteinmolekülen
anzufüllen. Darüberhinaus würde die Anzahl aller
Atome (etwa 1080) im Weltall, bei weitem nicht
ausreichen, um diese Vielfalt zu synthetisieren:
Durchmesser eines kugelförmigen Proteins von 200
Aminosäuren: ca. 5 nm; Volumen: ca. 65 nm3
Zahl aller möglichen Proteine mit 200 Aminosäuren: 1.267
x 10130
Gesamtvolumen von 10130 Proteinen: 9.7 x
1057 Kubiklichtjahre!!!
Zusammenfassung
Aufgrund der derzeit bekannten molekularbiologischen Grundlagen würde ich zur Biochemie und molekularen Genetik im Perryversum folgendes Basismodell vorschlagen:
Chiralität: Die Chiralität von biologischen Molekülen im Standarduniversum kann prinzipiell unterschiedlich sein. Lebensformen, die sich aus On und Noon Quanten entwickelt haben, weisen grundsätzlich eine homogene Chiralität für die sie kennzeichnenden biologischen Bausteinen auf. Bei Lebensformen, die sich früher entwickelt haben oder die sich generell ohne kosmokratischer Hilfestellung entwickelten, können bei ihren biologischen Bausteinen auch eine unterschiedliche Chiralität aufweisen.
Erbsubstanz: Die Erbsubstanz bei den Sauerstoffatmern im Perryversum besteht aus DNA. Diese DNA ist im einfachsten Fall aus den bekannten vier Nucleotiden Adenin, Guanin, Cytosin und Thymidin aufgebaut. Der benutzte genetische Code beruht ebenfalls auf Nucleotidtripletts (Codons), welche für die bekannten 20 Aminosäuren kodieren. Erweiterungen können wie folgt aussehen:
- Veränderte Nucleotide und veränderte oder zusätzliche Aminosäuren. Veränderte Aminosäuren wären sicherlich bei Oxtornern und anderen Extremwelt-Lebensformen angebracht.
- Erweiterte Anzahl von Nucleotide (6 Nucleotide) und daraus resultierender erweiterter genetischer Code, der aber gleichfalls auf Codons beruht. Damit einhergehend wäre eine deutliche Erweiterung der Anzahl der Aminosäuren.
- Unterschiedliche Realisierungen der Erbinformation. Statt DNA könnten Peptid-Nucleinsäuren (PNA) oder Pyranosyl-RNA (pRNA) verwendet werden.
Literatur
- Introduction to Protein structure. Carl Branden & John Tooze. Garland Publishing, Inc., 1991, ISBN 0-8153-0270-3
- Biochemie. Lubert Stryer. Spektrum der Wissenschaft Verlagsges. mbH, 1990, ISBN 3-89330-690-0
- Gene. Benjamin Lewin. VCH Verlagsges. mbH, 1991, ISBN 3-527-28052-9
- Protein Biosynthesis. H.R.V. Arnstein & R.A. Cox. Oxford Univ. Press, 1992, ISBN 0-19-963040-2
- The Hypercycle - A principle of natural self-organization. Manfred Eigen & Peter Schuster. Naturwissenschaften 1977, 64, 541-65.
